扁桃体って聞いたことあるでしょうか?側頭葉の内側の海馬の近くにある脳の部位で、アーモンド(扁桃)の形をしているので、このように呼ばれています。長らく情動・感情に関係があるといわれてきましたが、これは、てんかん治療で脳を取り除いたりしたときや、サルなどの除脳実験などで分かってきたことです。今回の論文では、このようなざっくりとした理解だった扁桃体の仕組みを最新の技術でかなり細かく調べています。将来的には、この論文で見られたニューロンを特異的に刺激してやることによって、好き嫌いも操作できるようになるかも知れません。
“Genetically identified amygdala-striatal circuits for valence-specific behaviors”
Xian Zhang et al., Nat Neurosci. 2021 Oct 18. doi: 10.1038/s41593-021-00927-0. Online ahead of print.
https://www.nature.com/articles/s41593-021-00927-0
・基底外側扁桃体(basolateral amygdala, BLA)のFezf2を発現するニューロンに、正または負の価数(感情価)の刺激に応答するニューロン(正価/負価ニューロン)がある。
・正価/負価ニューロンは、感覚領域と辺縁領域の別々のセットから入力を受け取り、嗅結節と側坐核への投影する。
・正価/負価ニューロンは、報酬と罰の情報をそれぞれ伝達して、正と負の強化を促進する。
<技術解説>
・Fezf2-CreERマウスについて
前回でも出てきた、Cre-LoxPシステムが使われています。CreはDNA組み換え酵素で、DNA上のloxP配列を認識して、DNAを特異的に組み換えます。これを利用して、特定の細胞において、特定の遺伝子をノックアウトする、コンディショナルノックアウト(Conditional Knock out)が良く使われます。
https://bsd.neuroinf.jp/wiki/Cre/loxP%E3%82%B7%E3%82%B9%E3%83%86%E3%83%A0
この論文では、Fexf2遺伝子を発現する細胞で、蛍光タンパク質やカルシウムプローブなどを発現させる用途に使われています。
・アデノ随伴ウイルス(AAV)について
AAVは、細胞の感染効率が高く、他のウイルスより非常に毒性が低いことから、研究用途だけでなく、遺伝子治療などにも応用されています。
https://www.cosmobio.co.jp/support/technology/a/adeno-associated-virus-aav-apb.asp
この研究では、特定の脳部位の神経を標識するためなどに使われています。
・GCaMP6fカルシウムイメージングについて
前回でも使われた技術で、神経活動をタンパク質のカルシウムプローブで調べることができます。神経活動が起こると細胞内のカルシウム濃度が高くなります。
・古典的条件付けについて
パブロフの犬の実験で、パブロフは無条件刺激(Unconditioned Stimulus, US)として、「餌」を使いました。この場合、無条件に応答として犬は唾液を出します。条件刺激(Conditioned Stimulus, CS)として、「ベル音」を使いました。ここで、CS>USを組み合わせて、繰り返し与えることで、CS(ベル音)だけで唾液が出るようになります。この研究では、CSを特定の音刺激、USを喉が渇いた時の「水」(吸水反応誘導)、「空気のパフ」(忌避反応誘導)を使いました。
・狂犬病ウイルスについて
狂犬病ウイルスは、神経細胞に感染して、神経から神経へと感染を広げる性質があり、これを利用して、神経回路を調べる実験に用いられます。
https://www.jnss.org/others/virus_vector/Rabies.htm
・DREDD、光遺伝学について
前回の論文でも出てきましたね。DREDDは薬理的に特定の神経活動を操作する方法。光遺伝学(Optogenetics)は、光刺激を使って特定の神経活動を操作する方法です。この論文では、Fezf2(BLa)神経の活動を選択的に操作しています。
<論文の流れ>
・BLa内のFezf2+細胞の同定
本研究で注目するFezf2+細胞を免疫組織染色で調べたところ、BLA内のFezf2+細胞は、多くはbasolateral nucleus (BLa)に存在し、 posterior basolateral nucleus (BLp)には少なかった(BLa, 93.5% ± 0.65%; BLp, 6.5% ± 0.65%)。この細胞は、 興奮性pyramidal neuronマーカーのCaMKII陽性だった。
・ Fezf2+細胞のGCaMP6カルシウムイメージング
BLaにおけるFezf2+細胞をGRINレンズで生きたまま活動を記録した。これにより、USとして水、air-puff 刺激を用いた。この結果から、正の感情価神経(positive-valence neurons, PVNs) と、 負の感情価神経(negative-valence neurons, NVNs)が、Fezf2+細胞に存在することを示した。
・active approach and avoidance (AAA)タスク(古典的条件付け)
特定の音刺激をCSとして、USと組み合わせることで、マウスを学習させた。トレーニングを繰り返すことにより、マウスは特定の音で、(CSR)>水吸引行動、または(CSP)>忌避行動を示すようになった。この時のFezf2(BLa)の活動を確認すると、CSRに応答する細胞、CSPに応答する細胞は増加した。
・条件付けと行動観察
AAAタスク中における行動と、Fezf2(BLa)神経の活動の相関関係を調べたところ、学習の過程と、神経活動はよく一致していた。神経活動から学習のスコア=動物の行動を推定できる。
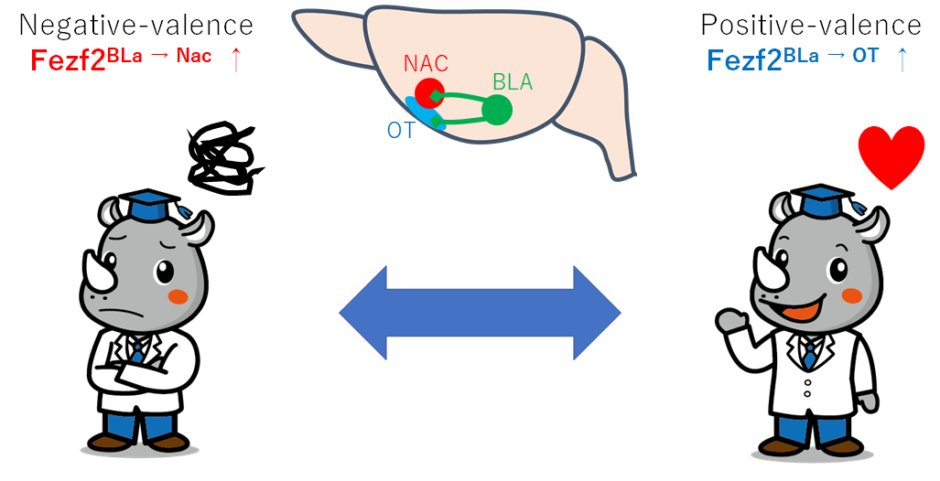
・Fezf2(BLa)神経の投射を調べた
AAVのプローブをBLaに注射することで、Fezf2(BLa)神経をGFPで標識することができる。これにより、全脳での投射を調べた。この結果、nucleus accumbens(NAc), olfactory tubercle (OT), medial prefrontal cortex(mPFC) and lateral septum (LS)が見いだされたが、特にNAc とOTが良く標識された。次に、NAc (Fezf2BLa → NAc)とOT (Fezf2BLa → OT)から逆行性に細胞を標識すると、BLA内ではほとんどが、Fezf2(BLa → NAc)神経(93% ± 2.5%)とFezf2(BLa → OT)神経 (92% ± 2.4%)であり、BLpでは少なかった。
・GCaMP6カルシウムイメージングでFezf2(BLa)神経投射経路を調べた
NAc (Fezf2BLa → NAc)とOT (Fezf2BLa → OT)の経路を逆行性でGCaMP6を発現させた。すると、NAc (Fezf2BLa → NAc)経路は、エアーパフ、電気ショックなど嫌悪刺激、OT (Fezf2BLa → OT)経路は、吸水など欲求刺激に応答した。逆はほぼ見られなかった。条件付けによって、これらの神経活動は強化された。
・Fezf2(BLa)神経への入力経路を調べた
NAc (Fezf2BLa → NAc)とOT (Fezf2BLa → OT)の経路に関わる入力を調べるために、BLaに狂犬病ウイルスを注入し、Fezf2(BLa)神経の上流の入力を調べたところ、NAc (Fezf2BLa → NAc)経路ではparaventricular nucleus of the thalamus (PVT), the cortical amygdala (CoA) and dorsal raphe (DR)が見られ、OT (Fezf2BLa → OT)経路では、piriform cortex (PIR) and the ventral part of the medial geniculate complex (MGv)が見られた。
・DREDDによるFezf2(BLa)神経活動の抑制
DREADD (designer receptor exclusively activated by designer drug)の技術を用いて、NAc (Fezf2BLa → NAc)とOT (Fezf2BLa → OT)の経路を選択的に抑制したところ、CSPによる忌避行動、CSRによる吸水行動、がそれぞれ抑制された。それぞれの他の行動には影響を示さなかった。
・光遺伝学GtACR2によるFezf2(BLa)神経活動の抑制
light-sensitive Guillardia theta anion-conducting channelrhodopsin 2 (GtACR2)を用いてNAc (Fezf2BLa → NAc)とOT (Fezf2BLa → OT)の経路を選択的に抑制したところ、CSPによる忌避行動、CSRによる吸水行動、がそれぞれ抑制された。それぞれの他の行動には影響を示さなかった。
・光遺伝学ChR2によるFezf2(BLa)神経活動の活性化
channelrhodopsin 2(ChR2)によって、NAc (Fezf2BLa → NAc)とOT (Fezf2BLa → OT)の経路を選択的に活性化したところ、real-time place preference or aversion (RTPP/RTPA) testにおいて、NAc経路刺激で、部屋を忌避し、OT経路刺激で部屋を嗜好することがわかった。また、two-port choice taskにおいては、OT経路刺激のスイッチを押し続けることがわかった。ローラーを使ったAAA taskにおいては、NAc経路刺激のみでCSによる忌避行動獲得ができることがわかった。
・脳の発達との関係が知りたい。ヒトでの好き嫌いの形成とのリンクはあるのか?
>脳の発生段階でどのくらいに回路が形成されるのだろうか?ヒトの好き嫌いとの関係はあるか?反抗期とか。
・Fezf2BLa neuronsのDREDDにより、記憶が無くなったといえるのか?反射の消失?
>マウスの記憶を調べるのは難しいが、記憶が無くなったのか?体が動いてしまうのが無くなったのか?=マウスでは区別できないのだろうか?
・Fezf2BLa neuronsのGtACR2抑制により覚えることができない
>ヒトで好き嫌いが激しい人とそうでもない人では、ここに違いがあったりするかな?
・Fezf2BLa neuronsのChR2刺激の結果は依存の行動を想起させる
>OT経路のtwo-port choice taskなどは、ドーパミン神経刺激の依存症モデルを想起させる。このモデルでドーパミンはどのように関連するだろうか?
・シナプス可塑性、強化はどこで起こっているのか?
>AAAタスクでは、Fezf2(BLa)神経活動が活性化している(すなわち上流が強化)。一方で、GtACR2抑制で、学習の獲得が阻害される(すなわち下流が強化?)。上流と下流のどちらで強化が起こっているか?(GtACR2抑制はフィードバックの関係かもしれない。)
基底外側扁桃体(BLA)は、正または負の価数(感情価)の刺激によって動機付けられた行動に不可欠な役割を果たしますが、動機付けに関して反対する情報を処理し、価数固有の行動の確立にどのように関与するかは不明です。ここでは、BLAでFezf2を発現するニューロンをターゲットにすることにより、動作中のマウスの2つの機能的に異なるクラス、それぞれ嫌悪刺激とやりがいのある刺激を本質的に表す負価ニューロンと正価ニューロンを識別して特徴づけます。罰の回避または報酬の追求に対応した、これら2つのクラスのニューロンは、感覚領域と辺縁領域の別々のセットから入力を受け取り、側坐核と嗅結節への投影を通じて罰と報酬の情報をそれぞれ伝達し、負と正の強化を促進します。したがって、感情価固有のBLAニューロンは、それぞれの入出力構造で配線され、感情価固有の動機付けられた動作のエンコード、学習、および実行におけるBLAの役割をサポートする回路フレームワークを形成します。
Xian Zhang et al., Nat Neurosci. 2021